813
Views & Citations10
Likes & Shares
The pathogenic yeasts of Cryptococcus neoformans species complex have been isolated in the environment from bird excreta, soil, trees and decaying wood. Although previous studies showed the ability of C. neoformans species complex to grow and mate on bird excreta and plant materials, there are no studies on soil. Therefore, the aim of the present study is to verify if C. neoformans species complex is able to grow on media containing soil and if it is able to sexually reproduce on this substrate. Soil media were prepared using four different soil samples collected in four different Italian regions, and the two reference strains JEC20 and JEC21 were tested to verify their ability to mate on these media. All the media supported the growth and sexual reproduction confirming that soil represents a realized niche for C. neoformans species complex.
Keywords: Cryptococcus neoformans, Soil, Sporulation, Yeasts, Murashige Skoog medium
INTRODUCTION
Encapsulated yeasts belonging to Cryptococcus neoformans and Cryptococcus gattii species complexes are the main fungal infectious agents of cryptococcosis, a life-threatening fungal infection occurring in both immunocompromised and immunocompetent hosts causing meningitis and meningo-encephalitis as the most frequent symptom [1]. The infection is acquired by inhalation of small cells or basidiospores present in the environment, which are able to reach pulmonary alveoli and to cause the onset of the disease [2]. Understanding the ecology of these fungal pathogens is a crucial step to elucidate which are their ecological niches and therefore the major sources and reservoir in the environment in order to prevent infection of hosts included in high risk categories [3]. In the environment, C. neoformans species complex was isolated from bird excreta, soil, trees, and decaying wood [4-8]. A recent environmental survey carried out in Europe showed also that some trees, such as olive trees and carob trees, present a higher rate of colonization than others. In addition, soil collected near the colonized trees was often contaminated with the fungus suggesting that trees are a potential reservoir and soil is a secondary niche [6]. Although previous studies showed the ability of C. neoformans species complex to grow and mate on bird excreta [9] and plant materials [10], there are no studies on soil. Therefore, the aim of the present study is to verify if C. neoformans species complex is able to grow on media containing soil and if it is able to sexually reproduce on this substrate.
MATERIALS AND METHODS
Sampling and soil medium preparation
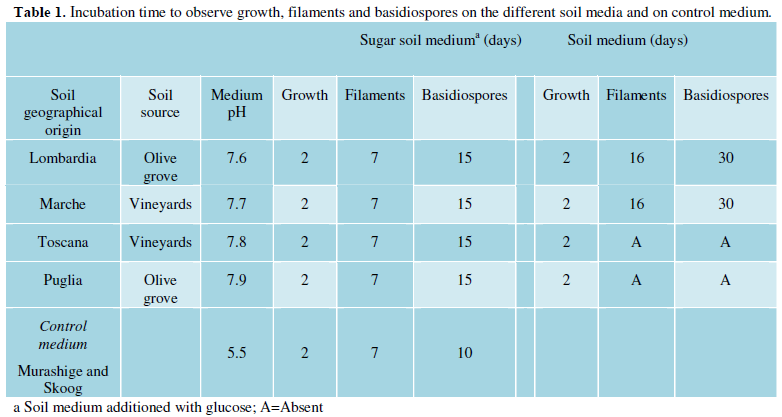
Soil samples were collected in four different regions of Italy: Lombardia, Toscana, Marche and Puglia (Figure 1). Soil samples from Marche and Toscana were collected from vineyards whereas those from Lombardia and Puglia were from olive groves (Table 1). Soil samples were not contaminated with bird excreta.
Soil medium (SM) was prepared dissolving 50 g of soil in 500 ml of distilled water. The suspension was boiled for at least 30 min and then filtered throughout some overlapping pieces of gauze. Volume was adjusted with further distilled water and 20 g of agar were added. The medium was sterilized by autoclave and then poured in 90 mm Petri dishes. For sugar soil medium (SSM), 2.5 g of glucose were added. Medium pH was measured before storage at 4°C. In addition Murashige Skoog medium (Sigma-Aldrich, Milano, Italy) was prepared as control mating medium.
Reference strains
Two reference strains were used in mating assays: JEC20 (Cryptococcus neoformans var. neoformans, mating type a) and JEC21 (Cryptococcus neoformans var. neoformans, mating type a). The strains were obtained by the National Institutes of Health (NIH), Bethesda, USA.
Mating assays
SM and SSM were prepared for each of the four soil samples. Each plate was inoculated with three streaks: one with JEC20 strain alone, one with JEC21 strain alone, and the third with the mix of the two strains. Plates were then incubated for four weeks at 25°C in the dark. Plates were observed periodically for the presence of filaments by a stereomicroscope (Zeiss, Oberkochen, Germany). Once the filaments appeared the culture was let grow for a further week and then a piece of agar containing hyphae was collected and observed by an optical microscope (Zeiss) in order to check and photograph the presence of clamp connections, basidia and basidiospores. One plate containing Murashige Skoog medium was also inoculated as described above in order to test if the two tester strains were actually able to mate.
RESULTS
The four media prepared with the four soil samples presented different pH values ranging from 7.6 to 7.9. Soil from Lombardia presented the lowest pH and that from Puglia the highest one (Table 1). The two reference strains were able to grow on all soil media with a visible growth after 48 h of incubation. On SSM agar, filaments become macroscopically visible at the edge of the streak of all mixed cultures at days 7 of incubation. The same streaks produced basidia and basidiospores at day 15. On SM, filaments and basidiospores were produced only on two mixed culture streaks (soil samples from Lombardia and Marche) at day 16 and 30, respectively (Table 1 and Figure 1). As expected streaks inoculated with JEC20 or JEC21 strains alone did not produce any filament in all plates. The two tester strains confirmed their ability to mate on Murashige and Skoog medium.
DISCUSSION AND CONCLUSION
This study shows that C. neoformans species complex is able to grow on media containing soil material either with or without addition of glucose. This means that soil is a suitable substrate for the growth and survival of this fungus in agreement with several studies reporting isolation of C. neoformans species complex from soil samples [6,7,11,12] Cryptococcus neoformans species complex has been shown to grow and to mate on medium containing pigeon excreta [9] as well as on medium containing plant materials [10] but, at present, similar studies on media containing soil has not yet been carried out. Our results show that soil represents a realized niche for C. neoformans species complex where it can grow and sexually reproduce. In addition, we showed that soil enables growth and mating of this yeast also without bird excreta contamination and confirming that Cryptococcus neoformans species complex is able to colonize a wide range of microhabitats. Potentially, each kind of soil is able to support sporulation of this pathogen especially if it is rich of carbohydrates and it is sufficiently acidic. This is also suggested by our results which showed that media prepared with soil from Puglia and Toscana, which were the least acidic media, enabled sporulation only in presence of glucose, whereas on the other two more acidic media (Lombardia and Marche) sporulation occurred also without glucose addition.
Basidiospores are the most resistant propagules of C. neoformans species complex and can be easily dispersed in the environment by wind, water, insects, birds, and other animals, as well as by human activities [13].
Therefore, the ability to produce basidiospores in soil represents an important step in Cryptococcus life cycle. In addition, presence of the fungus in the soil as yeast or basidiospores represents a potential source of infection because soil aerosols could transfer small cells and spores in pulmonary alveoli of humans and animals causing the onset of the infection as shown in previous studies [14,15]. At present, little is known about interaction between Cryptococcus neoformans and soil microhabitat, and many questions remain opened: how long can survive cryptococcal yeasts or basidiospores in soil? How do they interact with other organisms in this microhabitat? What is the abundance of this yeast in soil, and which kind of soil does better support their growth and mating? Further studies and efforts addressed to understand these and other questions about the ecology of this important fungal pathogen are needed in future.
1. Heitman J, Kozel T, Kwon-Chung KJ, Perfect J, Casadevall A (2010) Cryptococcus: From human pathogen to model yeast. American Society for Microbiology Press, Washington, DC, USA.
2. Lin X (2009) Cryptococcus neoformans: Morphogenesis, infection and evolution. Infect Genet Evol 9: 401-416.
3. Cogliati M, Puccianti E, Montagna MT, De Donno A, Susever S, et al. (2017) Fundamental niche prediction of the pathogenic yeasts Cryptococcus neoformans and Cryptococcus gattii in Europe. Environ Microbiol 19: 4318-4325.
4. Dou H, Wang H, Xie S, Chen X, Xu Z, Xu Y (2017) Molecular characterization of Cryptococcus neoformans isolated from the environment in Beijing, China. Med Mycol 55: 737-747.
5. Ellabib MS, Aboshkiwa MA, Husien WM, D'Amicis R, Cogliati M (2016) Isolation, identification and molecular typing of Cryptococcus neoformans from pigeon droppings and other environmental sources in Tripoli, Libya. Mycopathologa 181: 603-608.
6. Cogliati M, D'Amicis R, Zani A, Montagna MT, Caggiano G, et al. (2016) Environmental distribution of Cryptococcus neoformans and C. gattii around the Mediterranean basin. FEMS Yeast Res 16.
7. Escandón P, Castañeda E (2015) Long-term survival of Cryptococcus neoformans and Cryptococcus gattii in stored environmental samples from Colombia. Rev Iberoam Micol 32: 197-199.
8. Alvarez C, Barbosa GG, Oliveira R de V, Morales BP, Wanke B, et al. (2013) Techniques for the detection of pathogenic Cryptococcus species in wood decay substrata and the evaluation of viability in stored samples. Mem Inst Oswaldo Cruz 108: 126-129.
9. Nielsen K, De Obaldia AL, Heitman J (2007) Cryptococcus neoformans mates on pigeon guano: implications for the realized ecological niche and globalization. Eukaryote Cell 6: 949-959.
10. Springer DJ, Mohan R, Heitman J (2017) Plants promote mating and dispersal of the human pathogenic fungus Cryptococcus. PLoS One 12: e0171695.
11. Machado CC, Amaral AA, Severo LC (1993) Cryptococcus neoformans var. neoformans isolated from soil. Rev Inst Med Trop Sao Paulo 35: 77-79.
12. Montagna MT, Santacroce MP, Caggiano G, Tatò D, Ajello L (2003) Cavernicolous habitats harboring Cryptococcus neoformans: Results of a speleological survey in Apulia, Italy, 1999-2000. Med Mycol 41: 451-455.
13. Uejio CK, Mak S, Manangan A, Luber G, Bartlett KH (2015) Climatic influences on Cryptococcus gattii populations, Vancouver Island, Canada, 2002-2004. Emerg Infect Dis 21: 1989-1996.
14. Neilson JB, Fromtling RA, Bulmer GS (1977) Cryptococcus neoformans: Size range of infectious particles from aerosolized soil. Infect Immun 17: 634-638.
15. Ruiz A, Bulmer GS (1981) Particle size of air borne Cryptococcus neoformans in a tower. Appl Environ Microbiol 41: 1225-1229.
-
 Table 1
Table 1

QUICK LINKS
- SUBMIT MANUSCRIPT
- RECOMMEND THE JOURNAL
-
SUBSCRIBE FOR ALERTS
RELATED JOURNALS
- Journal of Genetics and Cell Biology (ISSN:2639-3360)
- Journal of Genomic Medicine and Pharmacogenomics (ISSN:2474-4670)
- Proteomics and Bioinformatics (ISSN:2641-7561)
- Journal of Astronomy and Space Research
- Journal of Biochemistry and Molecular Medicine (ISSN:2641-6948)
- Journal of Agriculture and Forest Meteorology Research (ISSN:2642-0449)
- Journal of Womens Health and Safety Research (ISSN:2577-1388)


